The Role of Local Backrub Motions in Evolved and Designed
Mutations,
by D. Keedy, I. Georgiev, E. Triplett, B. R. Donald, D. Richardson, and J. Richardson.
PLoS Computational Biology 2012;
8(8): e1002629. doi:10.1371/journal.pcbi.1002629
Author Summary.
Protein design has the potential to generate useful molecules for
medicine and chemistry, including sensors, drugs, and catalysts for
arbitrary reactions. When protein design is carried out starting from
an experimentally determined structure, as is often the case, one
important aspect to consider is backbone flexibility, because in
response to a mutation the backbone often must shift slightly to
reconcile the new sidechain with its environment. In principle, one
may model the backbone in many ways, but not all are physically
realistic or experimentally validated. Here we study the "backrub"
motion, which has been previously documented in atomic detail, but
only for sidechain movements within single structures. By a
two-pronged approach involving both structural bioinformatics and
computation with a principled design algorithm, we demonstrate that
backrubs are sufficient to explain the backbone differences between
mutation-related sets of very precisely defined motifs from the
protein structure database. Our findings illustrate that backrubs are
useful for describing evolutionary sequence change and, by extension,
suggest that they are also appropriate for rational protein design
calculations.
 Abstract.
Amino acid substitutions in protein structures often require subtle
backbone adjustments that are difficult to model in atomic detail. An
improved ability to predict realistic backbone changes in response to
engineered mutations would be of great utility for the blossoming
field of rational protein design. One model that has recently grown
in acceptance is the backrub motion, a low-energy dipeptide rotation
with single-peptide counter-rotations, that is coupled to dynamic
two-state sidechain rotamer jumps, as evidenced by alternate
conformations in very high-resolution crystal structures. It has been
speculated that backrubs may facilitate sequence changes equally well
as rotamer changes. However, backrub-induced shifts and experimental
uncertainty are of similar magnitude for backbone atoms in even
high-resolution structures, so comparison of wildtype-vs.-mutant
crystal structure pairs is not sufficient to directly link backrubs to
mutations. In this study, we use two alternative approaches that
bypass this limitation. First, we use a quality-filtered structure
database to aggregate many examples for precisely defined motifs with
single amino acid differences, and find that the effectively amplified
backbone differences closely resemble backrubs. Second, we directly
apply a provably-accurate, backrub-enabled protein design algorithm to
idealized versions of these motifs, and discover that the
lowest-energy computed models match the average-coordinate
experimental structures. These results support the hypothesis that
backrubs participate in natural protein evolution and validate their
continued use for design of synthetic proteins.
Abstract.
Amino acid substitutions in protein structures often require subtle
backbone adjustments that are difficult to model in atomic detail. An
improved ability to predict realistic backbone changes in response to
engineered mutations would be of great utility for the blossoming
field of rational protein design. One model that has recently grown
in acceptance is the backrub motion, a low-energy dipeptide rotation
with single-peptide counter-rotations, that is coupled to dynamic
two-state sidechain rotamer jumps, as evidenced by alternate
conformations in very high-resolution crystal structures. It has been
speculated that backrubs may facilitate sequence changes equally well
as rotamer changes. However, backrub-induced shifts and experimental
uncertainty are of similar magnitude for backbone atoms in even
high-resolution structures, so comparison of wildtype-vs.-mutant
crystal structure pairs is not sufficient to directly link backrubs to
mutations. In this study, we use two alternative approaches that
bypass this limitation. First, we use a quality-filtered structure
database to aggregate many examples for precisely defined motifs with
single amino acid differences, and find that the effectively amplified
backbone differences closely resemble backrubs. Second, we directly
apply a provably-accurate, backrub-enabled protein design algorithm to
idealized versions of these motifs, and discover that the
lowest-energy computed models match the average-coordinate
experimental structures. These results support the hypothesis that
backrubs participate in natural protein evolution and validate their
continued use for design of synthetic proteins.
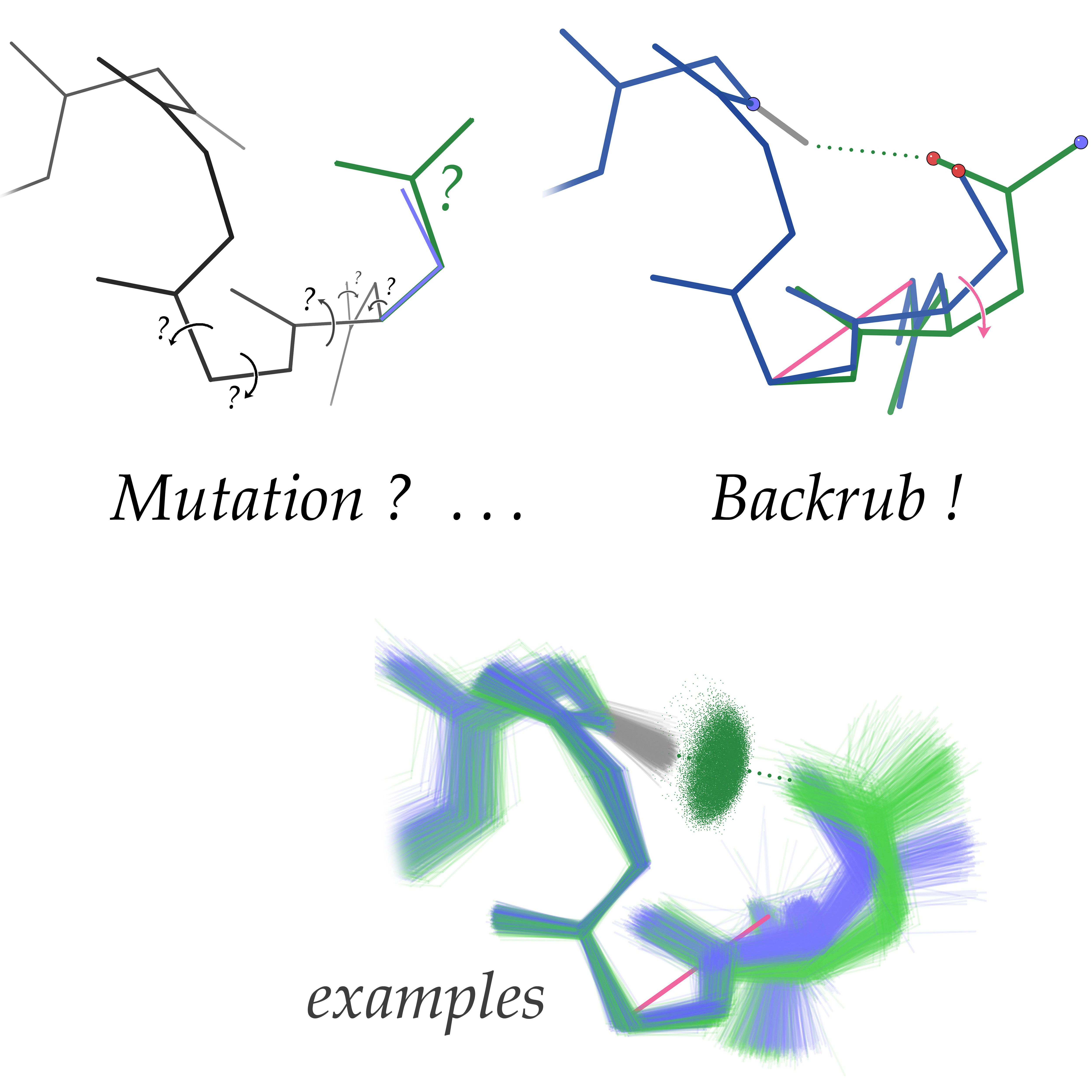
Figure:
The Ser->Asn helix N-cap mutation (top left) exemplifies cases that
require local backbone adjustment to enable favorable interactions
without steric clashes. Such backbone shifts involve complex
coordinated phi/psi rotations (shown) that are difficult to predict
accurately. However, the "backrub" motion can closely model many such
backbone changes with a single concerted movement, as computed for
this case by a deterministic, backrub-enabled protein design algorithm
(top right). Ensembles of Ser/Thr vs. Asn/Asp N-cap structural motifs
(bottom) increase the signal-to-noise ratio relative to single
wildtype-mutant structure pairs and support the conclusion that
backrubs accommodate actual amino-acid changes.
Read the free PDF.
 Abstract.
Amino acid substitutions in protein structures often require subtle
backbone adjustments that are difficult to model in atomic detail. An
improved ability to predict realistic backbone changes in response to
engineered mutations would be of great utility for the blossoming
field of rational protein design. One model that has recently grown
in acceptance is the backrub motion, a low-energy dipeptide rotation
with single-peptide counter-rotations, that is coupled to dynamic
two-state sidechain rotamer jumps, as evidenced by alternate
conformations in very high-resolution crystal structures. It has been
speculated that backrubs may facilitate sequence changes equally well
as rotamer changes. However, backrub-induced shifts and experimental
uncertainty are of similar magnitude for backbone atoms in even
high-resolution structures, so comparison of wildtype-vs.-mutant
crystal structure pairs is not sufficient to directly link backrubs to
mutations. In this study, we use two alternative approaches that
bypass this limitation. First, we use a quality-filtered structure
database to aggregate many examples for precisely defined motifs with
single amino acid differences, and find that the effectively amplified
backbone differences closely resemble backrubs. Second, we directly
apply a provably-accurate, backrub-enabled protein design algorithm to
idealized versions of these motifs, and discover that the
lowest-energy computed models match the average-coordinate
experimental structures. These results support the hypothesis that
backrubs participate in natural protein evolution and validate their
continued use for design of synthetic proteins.
Abstract.
Amino acid substitutions in protein structures often require subtle
backbone adjustments that are difficult to model in atomic detail. An
improved ability to predict realistic backbone changes in response to
engineered mutations would be of great utility for the blossoming
field of rational protein design. One model that has recently grown
in acceptance is the backrub motion, a low-energy dipeptide rotation
with single-peptide counter-rotations, that is coupled to dynamic
two-state sidechain rotamer jumps, as evidenced by alternate
conformations in very high-resolution crystal structures. It has been
speculated that backrubs may facilitate sequence changes equally well
as rotamer changes. However, backrub-induced shifts and experimental
uncertainty are of similar magnitude for backbone atoms in even
high-resolution structures, so comparison of wildtype-vs.-mutant
crystal structure pairs is not sufficient to directly link backrubs to
mutations. In this study, we use two alternative approaches that
bypass this limitation. First, we use a quality-filtered structure
database to aggregate many examples for precisely defined motifs with
single amino acid differences, and find that the effectively amplified
backbone differences closely resemble backrubs. Second, we directly
apply a provably-accurate, backrub-enabled protein design algorithm to
idealized versions of these motifs, and discover that the
lowest-energy computed models match the average-coordinate
experimental structures. These results support the hypothesis that
backrubs participate in natural protein evolution and validate their
continued use for design of synthetic proteins.